|
|
| Line 30: |
Line 30: |
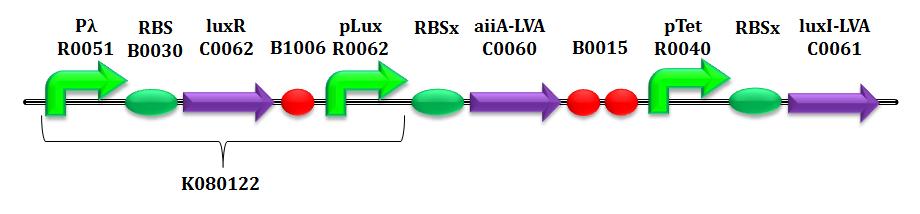
| | <b>CTRL-E.</b> is composed by two elements: a LuxI (BBa_C0061, 3OC6-HSL synthetase) expression cassette driven by the aTc-inducible pTet promoter and an AiiA (BBa_C0060, autoinducer lactonase) expression cassette driven by the 3OC6-HSL-inducible pLux promoter.</p> | | <b>CTRL-E.</b> is composed by two elements: a LuxI (BBa_C0061, 3OC6-HSL synthetase) expression cassette driven by the aTc-inducible pTet promoter and an AiiA (BBa_C0060, autoinducer lactonase) expression cassette driven by the 3OC6-HSL-inducible pLux promoter.</p> |
| | <p> | | <p> |
| - | In <em>E.coli</em> MGZ1 strain <a href="#Cox">(<i><b>Cox RS 3rd </b> et al. 2007</i>)</i></a>, singled out for the case study, pTet promoter is normally repressed, due to the presence of <em>tetR</em> gene, from Z1 cassette, integrated in its genome (<i><b>Lutz R</b> et al. 1997</i>): TetR product is able to inhibit the activity of pTet, thereby the 3OC6-HSL production. This allows the modulation of pTet activity by using tetracycline or anhydrotetracyclin (aTc) as inducers. A variation in the inducer concentration in input permits to modify the set-point of the 3OC6-HSL production in output. | + | In <em>E.coli</em> MGZ1 strain <a href="#Cox">(<i><b>Cox RS 3rd </b> et al. 2007</i>)</i></a>, singled out for the case study, pTet promoter is normally repressed, due to the presence of <em>tetR</em> gene, from Z1 cassette, integrated in its genome <a href="#"Lutz>(<i><b>Lutz R</b> et al. 1997</i>)</a>: TetR product is able to inhibit the activity of pTet, thereby the 3OC6-HSL production. This allows the modulation of pTet activity by using tetracycline or anhydrotetracyclin (aTc) as inducers. A variation in the inducer concentration in input permits to modify the set-point of the 3OC6-HSL production in output. |
| | When a critical amount of signal molecule is reached into the cells, the complex consisting of 3OC6-HSL and its transcriptional factor LuxR (constitutively expressed by pLambda promoter) is able to activate the pLux promoter, that regulates the expression of AiiA lactonase. | | When a critical amount of signal molecule is reached into the cells, the complex consisting of 3OC6-HSL and its transcriptional factor LuxR (constitutively expressed by pLambda promoter) is able to activate the pLux promoter, that regulates the expression of AiiA lactonase. |
| | So the HSL molecule regulates its own production via a negative feed-back loop system.</p> | | So the HSL molecule regulates its own production via a negative feed-back loop system.</p> |
| Line 48: |
Line 48: |
| | </table> | | </table> |
| | | | |
| - | <p>The circuit was built assembling <em>aiiA</em> protein generator and <em>luxI</em> translational unit with <partinfo>BBa_K081022</partinfo> composite part (<i><b>Pasotti L</b> et al. 2011</i>). Due to the length of the final circuit, the BioBrick parts were selected to reduce the internal homology, that could be cause of recombination or mutation events (<i><b>Sleight SC</b> et al. 2010</i>) (since the circuit is implemented in a strain expressing <em>recA</em> gene). The <partinfo>BBa_K081022</partinfo> part was purposely selected: in fact, the single terminator <partinfo>BBa_B1006</partinfo> and the double terminator <partinfo>BBa_B0015</partinfo> alignment does not show a significant sequence homology.</p> | + | <p>The circuit was built assembling <em>aiiA</em> protein generator and <em>luxI</em> translational unit with <partinfo>BBa_K081022</partinfo> composite part <a href="#Paso">(<i><b>Pasotti L</b> et al. 2011</i>)</a>. Due to the length of the final circuit, the BioBrick parts were selected to reduce the internal homology, that could be cause of recombination or mutation events <a href="#Sleight">(<i><b>Sleight SC</b> et al. 2010</i>)</a> (since the circuit is implemented in a strain expressing <em>recA</em> gene). The <partinfo>BBa_K081022</partinfo> part was purposely selected: in fact, the single terminator <partinfo>BBa_B1006</partinfo> and the double terminator <partinfo>BBa_B0015</partinfo> alignment does not show a significant sequence homology.</p> |
| | <p> | | <p> |
| | The circuit was designed without a terminator element downstream the <em>luxI</em> coding sequence. The lack of a terminator doesn't affect the behaviour of our circuit, since a terminator (<partinfo>BBa_B0054</partinfo>) is present in the low copy plasmid <partinfo>pSB4C5</partinfo> used. | | The circuit was designed without a terminator element downstream the <em>luxI</em> coding sequence. The lack of a terminator doesn't affect the behaviour of our circuit, since a terminator (<partinfo>BBa_B0054</partinfo>) is present in the low copy plasmid <partinfo>pSB4C5</partinfo> used. |
| - | The <em>aiiA</em> and <em>luxI</em> coding sequences are LVA tagged to decrease the protein half-life. (<i><b>Andersen JB</b> et al. 1998</i>)</p> | + | The <em>aiiA</em> and <em>luxI</em> coding sequences are LVA tagged to decrease the protein half-life. <a href="#Andersen">(<i><b>Andersen JB</b> et al. 1998</i>)</a></p> |
| | <p> | | <p> |
| - | In order to achieve the desired system output, a fine tuning of the whole circuit is required. A deeper understanding of the transcriptional and translational strength of the regulatory elements (promoter+RBS in several combination) and of the kinetic and the activity of the involved enzymes can be exploited to identify a mathematical model able to predict the behaviour of the controlled system, in order to avoid a cost and time expensive combinatorial approach.(<i><b>Howard M Salis</b> et al. 2009</i>)</p> | + | In order to achieve the desired system output, a fine tuning of the whole circuit is required. A deeper understanding of the transcriptional and translational strength of the regulatory elements (promoter+RBS in several combination) and of the kinetic and the activity of the involved enzymes can be exploited to identify a mathematical model able to predict the behaviour of the controlled system, in order to avoid a cost and time expensive combinatorial approach.<a href="#Salis">(<i><b>Howard M Salis</b> et al. 2009</i>)</a></p> |
| | <br> | | <br> |
| | </div> | | </div> |
| Line 131: |
Line 131: |
| | <div class="cleared"></div> | | <div class="cleared"></div> |
| | <div class="art-postcontent"> | | <div class="art-postcontent"> |
| - | </html>
| + | |
| | <div style='text-align:justify'> | | <div style='text-align:justify'> |
| | <ol type='1'> | | <ol type='1'> |
| - | <a name='Cox'><li>Cox RS 3rd, Surette MG, Elowitz MB (2007) <b> Programming gene expression with combinatorial promoters. </b> <i> Mol. Syst. Biol. </i> 3:145. </a> | + | <li><a name='Cox'></a>Cox RS 3rd, Surette MG, Elowitz MB (2007) <b> Programming gene expression with combinatorial promoters. </b> <i> Mol. Syst. Biol. </i> 3:145. </a> |
| | </li><br> | | </li><br> |
| - | <a name='Lutz'></a><li>Lutz R, Bujard H (1997) <b> Independent and tight regulation of transcriptional units in Escherichia coli via the LacR/O, the TetR/O and AraC/I1-I2 regulatory elements. </b> <i>Nucleic Acids Res.</i> 25(6):1203-10. | + | <li><a name='Lutz'></a>Lutz R, Bujard H (1997) <b> Independent and tight regulation of transcriptional units in Escherichia coli via the LacR/O, the TetR/O and AraC/I1-I2 regulatory elements. </b> <i>Nucleic Acids Res.</i> 25(6):1203-10. |
| | </li><br> | | </li><br> |
| - | <a name='Paso'></a><li>Pasotti L, Quattrocelli M, Galli D et al. (2011) <b>Multiplexing and demultiplexing logic functions for computing signal processing tasks in synthetic biology. </b> <i>Biotechnol. J. </i>6(7):784-95. </li><br> | + | <li><a name='Paso'></a>Pasotti L, Quattrocelli M, Galli D et al. (2011) <b>Multiplexing and demultiplexing logic functions for computing signal processing tasks in synthetic biology. </b> <i>Biotechnol. J. </i>6(7):784-95. </li><br> |
| - | <a name=''>Sleight</a><li>Sleight SC, Bartley BA, Lieviant JA et al. (2010) <b>Designing and engineering evolutionary robust genetic circuits. </b> <i>J. Biol. Eng. </i>4:12. | + | <a name=''Sleight'</a><li>Sleight SC, Bartley BA, Lieviant JA et al. (2010) <b>Designing and engineering evolutionary robust genetic circuits. </b> <i>J. Biol. Eng. </i>4:12. |
| | </li><br> | | </li><br> |
| - | <a name='Andersen'></a><li>Andersen JB, Sternberg C, Poulsen LK et al. (1998) <b>New unstable variants of green fluorescent protein for studies of transient gene expression in bacteria.</b> <i> Appl. Environ. Microbiol.</i> 64(6):2240-6. | + | <li><a name='Andersen'></a>Andersen JB, Sternberg C, Poulsen LK et al. (1998) <b>New unstable variants of green fluorescent protein for studies of transient gene expression in bacteria.</b> <i> Appl. Environ. Microbiol.</i> 64(6):2240-6. |
| | </li><br> | | </li><br> |
| - | <a name='Salis'></a><li>Salis HM, Mirsky EA, Voight CA (2009)<b> Automated design of synthetic ribosome binding sites to control protein expression. </b> <i>Nat. Biotechnol.</i>27:946-950. </li> | + | <li><a name='Salis'></a>Salis HM, Mirsky EA, Voight CA (2009)<b> Automated design of synthetic ribosome binding sites to control protein expression. </b> <i>Nat. Biotechnol.</i>27:946-950. </li> |
| | </ol> | | </ol> |
| | </div> | | </div> |
| | + | </html> |
| | {{end}} | | {{end}} |
 "
"